ومن المعروف أن عائلة موسعة من metazoan عامل ADP-ribosylation وADP-ribosylation عامل مثل GTPases صغيرة للعب أدوار أساسية في تحوير الوظائف الاتجار الغشاء وهيكل الخلية. هنا، نقدم التركيب البلوري للARL6، والطفرات التي تسبب متلازمة بارديه-بيدل (BBS3)، وتكشف مثل حلقة فريدة من نوعها التعريب في نهاية البعيدة الهيئات القاعدية، على مقربة من ما يسمى بوابة الهدبية حيث الحويصلات تحمل فتيل البضائع الهدبي مع الغشاء. الإفراط في GDP- أو أنواع من ARL6 / BBS3 GTP تخوض في الجسم الحي التأثيرات طول هدب الابتدائي وفرة. كما ينظم ARL6 / BBS3 يشير WNT، طريقا نقل الإشارة التي بالاشتراك مع أهداب في الفقاريات خرجت لتوها. الأهم من ذلك، يتم فقدان هذه الوظيفة في اشارة الى المتغيرات ARL6 تحتوي على الطفرات نقطة BBS المصاحب. من خلال تحديد هيكل متجهة الى GTP ARL6 / BBS3، إلى جانب المقايسات الفنية، ونحن نقدم التفسير الآلي لهذه التعديلات المسببة للأمراض، وهي غيرت النوكليوتيدات ملزمة. وبالتالي النتائج التي توصلنا إليها إنشاء دور لم تكن معروفة سابقا لARL6 / BBS3 في الهدبية الثدييات (ديس) تجميع وإشارات Wnt وتوفير المعلومات الأولى الهيكلية للبروتين BBS.
مقدمة
متلازمة بارديه-بيدل (BBS)
4 هو اضطراب البشري غير المتجانسة وراثيا تتميز مجموعة واسعة من الظواهر، بما في ذلك السمنة، وأمراض الكلى المتعدد الكيسات، تنكس الشبكية، polydactyly، والإعاقات الحسية
(1، -، 4). وهو يمثل نموذجا للciliopathies، فئة من الاضطرابات الجينية وتقاسم المسببات شيوعا من الجسم القاعدية و / أو اختلال وظيفي أهداب
(5، -، 9). أهمية الفسيولوجية للأهداب، والعضيات متحركة و / أو حسية موجودة على معظم أنواع الخلايا الفقاريات، وأكد من قبل عدد متزايد من الأمراض عديد المظاهر التي تنتمي إلى هذه الطبقة، التي بالإضافة إلى BBS تشمل متلازمة Alström ترتبط ارتباطا وثيقا، وكذلك متلازمة ميكل ومتلازمة جوبير، ومتلازمة العليا-لوكن.
فكر مرة واحدة ليكون عضية أثرية، والآن أنشأت امتحرك (الابتدائي) هدب فضلا عن كونها ضرورية لتنبيغ الكيميائية والبصرية، والمحفزات الحسية الميكانيكية، والغشاء مستقبلات رصع به ينسق عدد من أجهزة الطرد المركزي إشارات هامة وضرورية لحيوان فقاري التنمية، بما في ذلك القنفذ، WNT، وPDGFRαα الإشارات
(6، 10، -، 12).
يتم تجميعها الهدبية الخيط المحوري القائم على أنيبيب في النهاية البعيدة من الجسم القاعدية، بنية مشتقة من مريكز الأم التي الاحواض في غشاء البلازما قبل تكون الأهداب
(13، 14). تقريبا كلها مبنية أهداب وصيانتها من قبل وسائل النقل intraflagellar multiprotein (IFT) الآلية التي تحشد البضائع الهدبية، على سبيل المثال المكونات الهيكلية، والمستقبلات، وجزيئات الإشارة، في عضية عبر كينيسين السيارات تقدمي وينقل المكونات إلى قاعدة عن طريق محرك داينين الوراء
(15، -، 18). ويعتقد أن آلية IFT أن ترسو بالقرب من النصائح من الألياف الانتقالية، والتي هي هياكل التكوين معروف أن تشكيل-membered تسعة مجموعة شعاعي تشبه المروحة أن تنضم إلى النهاية البعيدة من الجسم القاعدية لغشاء الهدبية
(19، 20) . بالقرب من هذا الموقع، على نحو فعال "بوابة الهدبية،" الاتجار في مرحلة ما بعد جولجي من الشحنات حويصلي متجهة إلى هدب إنهاء الانصهار مع الغشاء. نقل الشحنات التي تطلق في عضية ويمكن بعد ذلك بتسهيل من الآلات IFT
(21، -، 23).
ومن المعروف أن عدد قليل من العوامل الخلوية للمشاركة في عملية التحول الوظيفي من الاتجار حويصلي للاتجار الهدبية
(24) أو ربما، في عملية عكسية حيث يتم الاتجار مكونات الهدبية من العضية. على الرغم من أن الاتجار الهدبي هو nonvesicular، فإنه يعتمد على IFT مفارز البروتين التي تشترك طبولوجيا بيتا المروحة / الملف اللولبي (والظاهر وظائف) مماثلة لتلك التي معطف البروتينات المرتبطة الحويصلة
(23، 25، 26). مجموعتنا اكتشف سابقا عضوا في الأسرة ARF تشبه من GTPases صغيرة، ARL6، التي يمكن أن امتلاك تصور هذه الوظيفة الاتجار. ويرتبط ARL6 مع بارديه-بيدل نوع متلازمة 3 (BBS3)
(27، 28)، مما يجعل الوقت الحاضر هو 1 من 14 البروتينات المرتبطة BBS
(4). في
انواع معينة ايليجانس أهداب الحسية، وقد تبين ARL6 للخضوع IFT
(28). وعلاوة على ذلك، والبروتينات ARF الشبيهة، والتي تورط عموما في الاتجار الغشاء وتنظيم العمليات المرتبطة أنيبيب
(29، -، 32)، واثنين من أعضاء على الأقل أن المشهودة وظائف الهدبية. orthologues ARL3 من
الليشمانيا، C. ترتبط
ايليجانس، والفئران مع وظائف أهداب / الهدبية مستقبلة للضوء
(33، -، 35)؛ وبالمثل، ARL13B يموضع خصيصا لأهداب في
C. ايليجانس، والماوس أنها تشارك في تشكيل هدب والقنفذ الإشارات
(34، 36). وجود الحصري لARL6 في الكائنات الحية التي تمتلك الأهداب وارتباطه IFT
(28 ول) يشير إلى وجود دور محتمل لهذا GTPase صغير في تنسيق أو تسهيل سد الوظيفي الاتجار intraciliary مع الاتجار حويصلي.
على الرغم من أن جميع البروتينات BBS درس حتى الآن يبدو أن تلعب دورا هاما في أي تهريب البضائع إلى القاعدية الجسم / الجسيم المركزي أو خلال أهداب عبر IFT
(2)، ويقترح ARL6 / BBS3 أن يكون لها وظيفة يحتمل أن تكون متميزة عن أن مجموعة أساسية من BBS البروتينات التي تجمع في مجمع، وصفه BBSome
(37). وتنشأ هذه الفكرة من هذا الاكتشاف أن BBSome، التي تتكون من بروتين معقد فصل كيميائيا تحتوي على BBS1، BBS2، BBS4، BBS5، BBS7، BBS8، والبروتينات BBS9، لا تشارك في يجزئ مع BBS3 في التدرج السكروز
(37). ومن المثير للاهتمام، يتفاعل عنصر واحد من BBSome (BBS1) مع Rabin8، وهو عامل الصرف الناتج المحلي الإجمالي / GTP لRAB8 GTPase صغيرة. Rabin8 المشارك يموضع مع BBSome في الجسيم المركزي / الجسم القاعدية ويؤثر على الدراجات النوكليوتيدات من RAB8، الذي يدخل هدب في ولاية محددة GTP لها وضروري لتكون الأهداب
(37). كما تبين RAB8 مؤخرا للتفاعل مع Rabaptin5، الذي يموضع في الجسم القاعدية، والزميلة مع البروتين IFT يطلق Elipsa / DYF-11
(38، 39). نظرا للدور راسخة من Rabaptin5 باعتباره المستجيب RAB5 في الإلتقام، واقترح أن هذا Rabaptin5-RAB8-Elipsa / DYF-11 الثلاثي قد توفر التفاعلات الحيوية الوظيفية بين الغشاء الهدبي (أو الحويصلات المرتبطة أهداب) وآلية IFT (
39). في
C. ايليجانس، RAB-5 وعرض نفسه للتركيز في قاعدة الأهداب إلى جانب مجمع المرتبطة الاندوسوم أوائل STAM-ساعة الذي ينظم إزالة polycystin-1 / polycystin-2 البروتين من أهداب
(40). كيف BBSome البروتينات، التي هي في حد ذاتها المرتبطة مع كل من الهيئة القاعدية وIFT، المشاركة في هذه العملية الفرز الاتجار الجسم الهدبي القاعدية لا يزال بعيد المنال، ودور BBS3، يحتمل أن تكون متميزة عن مكونات BBSome الأساسية، غير معروف في الوقت الحاضر.
لتسليط الضوء على وظائف الجزيئية والخلوية من ARL6 / BBS3، توصلنا لأول مرة التركيب البلوري للشكل محدد GTP من البروتين البشري. هيكل ARL6 / BBS3 يمثل أول من حلها لBBS أو البروتين IFT ويعرض خصائص فريدة لا توجد في GTPases الصغيرة الأخرى. في تركيبة مع GTP ملزمة المقايسات الفنية، يوضح هيكل كيف احد بقايا الطفرات كشفت في المرضى BBS إلغاء النوكليوتيدات ملزمة وبالتالي وظيفة البروتين. وعلاوة على ذلك، علينا أن نظهر الآن أن ARL6 في خلايا الثدييات المهدبة يموضع إلى الهيئات القاعدية النهاية البعيدة، قرب أو على الألياف التي تمر بمرحلة انتقالية، وبالتالي على مقربة من موقع حويصلة لرسو السفن / الانصهار في قاعدة الأهداب. وانسجاما مع دور للARL6 في هذا الموقع التحت خلوية، وتبين لنا أن يعطل وظيفتها الخلوية، عن طريق الإفراط في GDP- أو أشكال غير الساحلية GTP، يؤدي إلى الشذوذ الهدبية
في الجسم الحي. وعلاوة على ذلك، الزائد الخلوي من النوع البري ARL6 يسبب التنظيم تتكون من إشارات Wnt. الأهم من ذلك، هو لم يلاحظ هذا التأثير مع المتغيرات المسببة للأمراض من ARL6 / BBS3. معا، وتكشف النتائج التي توصلنا إليها أن بروتين BBS ليس مباشرة (أو بإحكام) المرتبطة BBSome يموضع إلى موقع الرواية في المنطقة الجسم القاعدية، حيث يمكن أن تنظم أهداب (ديس) تجميع وربما نتيجة لذلك، الهدبية يشير أحداث لاحقة . ونقترح أن ARL6 / BBS3 قد تؤدي هذا الدور عن طريق تحوير الاتجار الغشاء عند قاعدة العضية الهدبية.
للدراسات البلورات، وبقايا الأحماض الأمينية 16-186 من ARL6 من استنساخ الثدييات الجينوم مجموعة (الإحداثيات Geneservice AT30-A7) تم تضخيمها من قبل PCR وsubcloned في ناقلات pET28a-LIC (GI: 145307000)، وذلك باستخدام و-فيوجن PCR نظام الاستنساخ (Clontech). تم تحويل بناء إلى سلالة القولونية BL21-CodonPlus (DE3) -RIL (Stratagene)، وكانت تزرع الخلايا عند 37 درجة مئوية في رائع مرق إلى A 600 3.0 باستخدام نظام محتدما LEX. تم تخفيض حمام الماء إلى 15 درجة مئوية لمدة 30 دقيقة قبل الاستقراء باستخدام 1 م م الآيزوبروبيل 1-ثيو-β- د -galactopyranoside وثم غادرت عند 15 درجة مئوية خلال الليل. حصاد، تم طرد الخلايا، في النيتروجين السائل، وتخزينها في -80 ° C-المجمدة فلاش. ومعلق الكريات الخلايا إذابة من 1.8 لتر من ثقافة في 100 مل من تحلل العازلة (10 م م تريس، حمض الهيدروكلوريك، ودرجة الحموضة 7.5، 0.5 متر كلوريد الصوديوم، 5 م م إميدازول) مع 1 م م phenylmethylsulfonyl الفلوريد، و 0.5٪ CHAPS و lysed مع صدر microfluidizer في 20000 رطل مسح طاف من خلال عمود تبادل أنيون DE52 (هتما) معايرتها قبل مع تحلل العازلة ومن ثم تحميلها على 5 مل HisTrap HP (GE للرعاية الصحية) عمود أيضا قبل معايرتها مع العازلة تحلل. ومزال البروتين الهدف مع التدرج إميدازول 50-500 م م في 50 مل. تم جمع الكسور بروتين مزال، تجميع، وتنقيته من خلال تبني الترشيح هلام (Superdex 75) العمود معايرتها قبل مع العازلة التي تحتوي على 20 م م HEPES، ودرجة الحموضة 8.0، 500 م م كلوريد الصوديوم، و 1 م م dithiothreitol. تم تعديل الكسور البروتين التي تم جمعها لاحتواء 5 م م MgCl 2 ثم حضنت مع 5 × بروتين تركيز المولية من الناتج المحلي الإجمالي لمدة 1 ساعة. ثم تركزت البروتين باستخدام الترا Amicon 15 فلتر الطرد المركزي لتركيز النهائي من 79.2 ملغ / مل. كان البروتين النقي كتلة من 21،354.8 دا.
بلورة وتحديد الهيكل التنظيمي
وقد نمت بلورات الانعراج باستخدام يجلس انخفاض التبخر في درجة حرارة الغرفة. احتوى المحلول الام 2 م (NH 4) 2 SO 4، 0،2 م كلوريد الصوديوم، 0.1 م كاكوديلات الصوديوم، ودرجة الحموضة 5.5. يبدو الكريستال ركوب بعد 2 أيام. تم جمع البيانات حيود في النحاس الدورية مصدر الأنود (RIGAKU FR-E) ومعالجتها باستخدام HKL2000
(41) برمجيات. تم إجراء استبدال الجزيئي باستخدام PHASER البرنامج
(42) مع فرقة من الهياكل اثنين (رموز PDB
1ZD9 و2H16) كنموذج البحث. تم بناء نموذج ARL6 الأولي تلقائيا في ARP / السدى
(43)، تليها عدة جولات من ضبط النفس تنسيق وعامل درجة الحرارة التحسينات في REFMAC
(44)، والمصادقة الهندسة في MOLPROBITY
(45 الخادم)، ودليل التكيف في الطير المائي
(46). وتلخيص البيانات الحيود والإحصاءات نموذج في
الجدول 1. وقد تم إيداع الإحداثيات وعامل سعة هيكل في PDB كرمز
2H57 (47). البرية من نوع
الإنسان العاقل ARL6 كدنا] (بنك الجينات TM عدد الانضمام
NM_032146 تم استنساخ) في pRSET-6A
(48). وقد أعدت كل المسوخ ARL6 بطريقة الطفرات موقع الموجه المعدلة المعروفة باسم SLIM
(49). للحصول على البروتين GST الموسومة، تم استنساخ كامل طول ARL6 إلى pGEX-6P-1 (أمرشام العلوم البيولوجية).
واستخدمت BL21 (DE3) خلايا pLysS للتعبير عن غير محدد (pRSET) أو الموسومة GST (pGEX) من النوع البري وARL6 متحولة. تحولت E. وكانت القولونية الخلايا المستزرعة في 50 مل من مرق LB التي تحتوي على 34 ميكروغرام / مل الكلورامفينيكول و 50 ميكروغرام / مل الأمبيسلين عند 37 درجة مئوية حتى
A 600 = 0.6. استميلت الثقافات مع 0.4 م م الآيزوبروبيل 1-ثيو-β- بين عشية وضحاها -galactopyranoside د في درجة حرارة الغرفة. إلى الحصاد، ومكعبات الخلايا بواسطة الطرد المركزي في 5000 ×
ز لمدة 10 دقيقة، وعلقت في غسل العازلة (50 م م ناه 2 PO 4، 2 م م dithiothreitol، 0.3٪ توين 20، 4 μ م ATP)، و lysed صوتنة. تم فصل الأجزاء القابلة للذوبان وغير القابلة للذوبان عن طريق الطرد المركزي في 20000 ×
ز لمدة 10 دقيقة. وقد أجريت فحوصات ملزمة GTP كما هو موضح سابقا
(50، 51). لفحص الكسر القابلة للذوبان، ورصدت 10 ميكرولتر من المحللة على غشاء النيتروسليلوز، الذي تم غسلها بعد ذلك في غسل العازلة وسمح لاحتضان لمدة 2 ساعة في درجة حرارة الغرفة العازلة في ملزمة (50 م م ناه 2 PO 4، 2 م م dithiothreitol، 0.3٪ توين 20، 4 μ م ATP، 2.5 م م MgCl 2) مع 2 μCi من GTP / مل. التالي ملزمة، وتشطف الغشاء في غسل العازلة ويتعرض لتصوير أو شاشة الفوسفور. لفحص جزء بيليه، علقت البروتين في Laemmli عينة العازلة ومفصولة SDS-PAGE. وغارقة هلام في 50 م م تريس، حمض الهيدروكلوريك، ودرجة الحموضة 7.5، مع 20٪ الجلسرين لمدة 30 دقيقة في درجة حرارة الغرفة، ونقل السائل على نتروسليلوز. وقد تم TLC كما هو موضح سابقا
(50). النسر الأساسية الدنيا والمتوسطة
(إينفيتروجن كان يستخدم) للحفاظ على HEK-293 الخلايا، و1: خليط 1 من المعدل المتوسط Dulbecco والنسر وهام في F-12 المتوسطة
(إينفيتروجن كانت تستخدم) لزراعة الخلايا IMCD3 وhTERT-RPE. واستكملت كلا النوعين من وسائل الإعلام مع 10٪ مصل بقري جنيني، وكانت تزرع جميع الخلايا عند 37 درجة مئوية في 5٪ CO 2. لعلاج المخدرات، علقت نوكودازول في DMSO وتستخدم بتركيز نهائي من 0،1 ميكروغرام / مل لمدة 1 ساعة أو طوال الليل. كان
MYC -tagged بناء P50 dynamitin التعبير هدية عينية من ريتشارد فالي (جامعة كولومبيا) وكان transfected في الخلايا IMCD3 باستخدام Lipofectamine2000
(إينفيتروجن) وفقا لبروتوكول الشركة المصنعة، وكذلك كل البنى الأخرى المستخدمة.
لجميع التجارب مناعية، كانت تزرع الخلايا على coverslips الزجاج وتشطف مع الفوسفات مخزنة المالحة قبل التثبيت. تم إصلاح الخلايا إما عن طريق الحضانة 10 دقيقة في -20 درجة مئوية الميثانول، تليها الإماهة 10 دقيقة في TBS-T (20 م م تريس قاعدة، ودرجة الحموضة 7.5، 150 م م كلوريد الصوديوم، 0.05٪ توين 20)، أو عن طريق تثبيت 30 دقيقة مع 2٪ امتصاص العرق في PHEM (0.05 م أنابيب، 0.025 م HEPES، 0.01 م EGTA، 0.01 م MgCl 2، ودرجة الحموضة 7.2)، تليها استخراج 2-مين مع 0.1٪ تريتون X-100 في PHEM. بعد التثبيت، وخلايا منعت إما لمدة 1 ساعة في TBS-T تستكمل مع 1٪ البقري ألبومين المصل قبل الحضانة الأجسام المضادة الأولية أو المحتضنة مباشرة لمدة 60 دقيقة في درجة حرارة الغرفة مع الأجسام المضادة الأولية ذات الصلة مخففة في TBS-T. ثم تم غسلها الخلايا في TBS-T وحضنت 60 دقيقة أخرى في الضد الثانوية المناسب. تم counterstained نوى التي يحتضنها coverslips الزجاج في 100 ن م 4، 6-diamidino-2-phenylindole لمدة 5 دقائق في TBS-T في درجة حرارة الغرفة. وقد شنت الشرائح في إطالة مكافحة تتلاشى كاشف
(المسابر الجزيئية) والتي لاحظها مضان المجهر باستخدام epifluorescence القياسي، لايكا LSM 410 مبائر، أو لايكا TCS أنظمة المجهر SP متحد البؤر. أجريت مناعي المجهري متحد البؤر الرقمي وdeconvolution كما هو موضح سابقا
(52).
تم تنفيذ المناعية للأنسجة الخصية الماوس على الماوس جزءا لا يتجزأ من أكتوبر وسريعة المجمدة قبل باجتزاء. تم الحصول على أقسام الكلى الماوس من INOVA تشخيص، سان دييغو (الكتالوج رقم 508385)، والتي تم الحصول عليها قرد أقسام الدماغ من ميديكا، إنسينيتاس، CA (الكتالوج رقم 0608-BR). وقد أجريت تلطيخ والتصوير كما هو موضح أعلاه لخلايا ثقافة أنسجة الثدييات.
وتشمل الأجسام المضادة الأولية المستخدمة في مناعية مضادة للARL6C ومضادة للARL6N (Abgent)، ومكافحة γ تويولين (سيغما)، ومكافحة الأسيتيل تويولين (سيغما)، ومكافحة α تويولين (سيغما)، ومكافحة PCM1 (هدية من الدكتور أندرياس Merdes، جامعة ادنبره). وتشمل الأجسام المضادة الثانوية المستخدمة الأرنب والفأر مفتش مترافق مع فلور اليكسا 488 و 594
(المسابر الجزيئية). تم حساب الاختلافات في طول أهداب باستخدام ر اختبار الطالب، في حين تم حساب الاختلافات في النسبة المئوية للخلايا مع أهداب باستخدام اختبار χ 2.
WNT luciferase المراسل فحوصات
خلايا كلوة-293T معربا عن ثابت مراسل pTOPflash
(53 كانت المصنفة) والخلايا hTERT-RPE في مناطق ذات كثافة من 10 4 خلايا لكل بئر في لوحة 24-جيدا. بعد 24 ساعة، و transfected الخلايا مع ما مجموعه 1 ميكروغرام من الحمض النووي
(Renilla السيطرة luciferase المراسل، البلازميد من الاهتمام، ومراسل pTOPflash البلازميد عند الضرورة) باستخدام FuGENE 6 (روش العلوم التطبيقية) وفقا لتعليمات الشركة الصانعة. تم استنساخ BBS3 من النوع البري وأليل متحولة إلى pCMV وpcDNA 6.2 بروتين الفلورية الخضراء العمود الفقري واختبارها مع ناقلات الرقابة المناسبة على التوالي. وقد حفز الخلايا مع Wnt3a المتوسطة مكيفة
(54) 48 ساعة بعد ترنسفكأيشن وتحصد 24 ساعة بعد التحفيز. تم قياس luciferase النشاط باستخدام طقم المزدوج luciferase المراسل مراسل الفحص (PROMEGA) على TECAN Genios برو luminometer وBMG LABTECH FLUOstar أوميغا لوحة القارئ.
لإلقاء الضوء على وظيفة الجزيئية للARL6 / BBS3 وتسهيل الدراسات هيكل وظيفة، توصلنا إلى هيكل عالية الدقة من هذا GTPase صغير. باستخدام طريقة الجلوس انخفاض التبخر القياسية، حصلنا على بلورات الانعراج على ما يصل الى 2 قرار Å لشكل اقتطاع N-محطة من ARL6 (بقايا 16-186) في ولاية محددة GTP (ARL6-GTP)
(الجدول 1). ثم تم حلها البنية البلورية للبروتين عن طريق استبدال الجزيئي، كما هو موضح في إطار "إجراءات تجريبية".
الجدول 1
بيانات حيود والإحصاءات نموذج البلورات
rmsd، جذر متوسط الانحراف مربع.
هيكل مجمع ARL6-BBS3-GTP يظهر GTPase نموذجي أضعاف مع ورقة β المركزية الذين تقطعت بهم السبل ستة تحيط بها خمس سنوات اللوالب ألفا
(الشكل 1 A)، لكنه يختلف عن معظم GTPases الصغيرة التي يتحلل عادة GTP المربوطة وغالبا ما توجد في ولاية محددة الناتج المحلي الإجمالي. وعلى الرغم من التشابه بين ARL6-GTP إلى أخرى رأس GTPases الصغيرة الفصيلة في ولايات محددة التماثلية GTP أو GTP nonhydrolyzable، بما في ذلك التشكل من خمس الحفاظ G-مربع (G1-5) تسلسل
(55)، وثلاثة عناصر الهيكلية للARL6-GTP ، لا سيما في التبديل الأول، التبديل الثاني، والمناطق interswitch، وأظهرت الخلافات بتكوين مثيرة من ذلك من ARF1 في ولاية محددة الناتج المحلي الإجمالي (ARF1 الناتج المحلي الإجمالي)
(الشكل 1 B). اعتمدت منطقة التبديل لي التشكل الموسعة غير النظامية التي تنسجم مع التقيد GTP ولم تشكل β حبلا إلى الزوج مع β3 كما هو الحال في ARF1 الناتج المحلي الإجمالي
(الشكل 1 B). المنطقة I التحول من ARL6 هي اثنين من مخلفات أطول من أعضاء نموذجي آخرين من العائلة ARF، وتشكيل 3 10 -helix في بقايا 40-42
(الشكل 1 B). مجموعة أميد من الغليسين-72 في منطقة التبديل II تشكل السندات الهيدروجين مع مجموعة γ-فوسفات GTP ويتم نقل بزيادة 7.5 Å بالمقارنة مع المقابلة بقايا الغليسين-70 من ARF1 الناتج المحلي الإجمالي
(الشكل 1 C)؛ هذا التغيير مهم المحتمل للربط تفضيلية من GTP. في المطابقة لحركة بقايا التبديل الثاني الغليسين-72، وβ-دبوس الشعر من المنطقة interswitch (خيوط β2-β3) ويمتد ومعرضة للمذيب
(56). ومن المثير للاهتمام، ARL6 هو العضو الوحيد في GTPases أسرة المنتدى والصغيرة التي لديها سيرين (بقايا سر-71) في المركز الثاني من
X عزر D
XX G للتبديل المنطقة الثانية؛ كل GTPases ARF الشبيهة الأخرى لديها الجلايسين في نفس الموقف
(57). اقترحت مجموعة الهيدروكسيل سلسلة من الجانب سر-71 سندات أشكال الهيدروجين مع مجموعة أميد من GLN-73 وجزيء الماء لمهاجمة جماعة γ الفوسفات من GTP خلال التحلل
(الشكل 2 A). وتمشيا مع انخفاض درجة الحرارة عامل توزيع ARL6-GTP
(الشكل 1 D)، وهذه الروابط الهيدروجينية إضافية تجعل الشبكة بالكامل التفاعل التي تنطوي على المغنيسيوم 2+ وGTP، وخصوصا التبديل الأول والتبديل II المناطق، أكثر جمودا مقارنة GTP- الآخر GTPases المنتدى الاقليمى للاسيان ملزمة. من ناحية أخرى، وحلقة ربط β2 وβ3 أكثر مرونة مقارنة GTPases ARF الأخرى
(الشكل 1 D).

الشكل 1.
التركيب البلوري للARL6 / BBS3 GTPase صغير. A، تمثيل
اسطوانة من هيكل ARL6 في شكل محدد GTP لها. يتم تلوين عناصر الهيكل التي تخضع لتغييرات جوهرية على دورة النقد الناتج المحلي الإجمالي / GTP
الأرجواني لتبديل I، II
الأزرق للتبديل،
والأحمر للinterswitch. يتم عرض حلقة P
باللون الأصفر، ويظهر GTP ملزمة في
التمثيل عصا. B، مقارنة بنية محددة GTP ARL6 GTPase (رمز PDB
2H57) مع الالتزام الناتج المحلي الإجمالي كامل طول ARF1 GTPase (PDB كود
1HUR). عناصر الهيكل التي تمر بمرحلة تغير التشكل الأساسي في دورة الصرف الناتج المحلي الإجمالي / GTP، والتبديل الأول، التبديل الثاني، وinterswitch، واللون
الأرجواني، والأزرق، والأحمر للARL6
والسماوي والأزرق البنفسجي، والشوكولاته الأحمر في ARF1. يتم عرض جميع عناصر البنية الثانوية الأخرى من ARL6 في الشريط
(القمح)، على الرغم من ARF1 يظهر فقط Cα التتبع
(الخضراء). C، تغيير متعلق بتكوين الرئيسي في المنطقة التبديل الثاني عند تغيير الدول النوكليوتيدات المربوطة بين ARL6 (محددة GTP) و ARF1 (متجهة إلى الناتج المحلي الإجمالي). نظام الألوان هو نفسه كما هو الحال في
A. 70 الغليسين من ARF1 تظهر ذرات Cα من الغليسين-72 من ARL6 والمقابلة كما
المجالات. D، والمرونة التشكل الإقليمية محددة GTP ARF1
(يسار، PDB كود
1O3Y) مقارنة مع تلك التي منضم GTP ARL6 (رمز PDB
الصحيح 2H57). يتم تلوين شرائط وفقا
لB -factor قريب من ذرات Cα، حيث يتم تلوين ذرات Cα مع أعلى
B -factor
الأحمر، ويتم تلوين ذرات Cα مع أدنى
B -factor
الأزرق.

الشكل 2.
التركيب البلوري للARL6 / BBS3 يسلط الضوء على الإمراضية من المتغيرات وجدت في المرضى الذين يعانون BBS. A، عرض ستيريو شبكة التفاعل حول موقع نشط من GTPase صغير ARL6. وتظهر جزيئات الماء في 2+ أيون الأزرق والمغنيسيوم في البرتقال. وتظهر التفاعلات القطبية تتعامل مع الهجوم على جزيء الماء وسر-71 تقدم اللاعب خطوط حمراء. بقايا الثريونين-31، تحور الذي يسبب BBS، هو المسمى في أرجواني. B، عرض ستيريو لشبكة الرابطة الهيدروجينية حول صناديق G4 و G5 وكذلك الغليسين-169 من ARL6. وتظهر السلاسل الجانبية ذات الصلة ARL6 في التمثيل العصا، وتظهر العصي لGTP مع دائرة نصف قطرها أرق. وصفت الروابط الهيدروجينية التي تنطوي على جزيء GTP كخطوط متقطع السوداء. وصفت الروابط الهيدروجينية التي تنطوي على الغليسين-169 باللون الأحمر، والسندات الآخرين الهيدروجين التي تنطوي على بقايا لG4 و G5 هي باللون الأصفر. C، الأساسية مسعور حول لوي-170 من ARL6. وتظهر السلاسل الجانبية من لوي-37، إيل 33، الفنيل ألانين-128، ولوي-170 كما المجالات.
حل التركيب البلوري للARL6 / BBS3 وليس فقط سمح لنا لتوصيف بنية البروتين من النوع البري، ولكنها قدمت أفكارا بالحالة المرضية للبدائل حمض أميني واحد (T31M، T31R، G169A، وL170W) لدينا وذكرت المجموعة أن تسبب BBS في المرضى
(28). عدة مجموعات الأميد في مربع G1 (GX4GKT)، المعروف أيضا باسم P-حلقة، وتشكل روابط هيدروجينية الحاسمة مع triphosphates من النوكليوتيدات GTP. على وجه الخصوص، تساهم المجموعة سلسلة الهيدروكسيل منتدى المجالس الرومانسية-31 الجانب للتنسيق من المغنيسيوم 2+، β- وγ الفوسفات من GTP، جزيء الماء مجانا، وجزيء الماء سد لآسيا والمحيط الهادئ-69، عن أهمية حاسمة للنشاط GTPase من ARL6
(الشكل 2 A). فمن الممكن أن تحور منتدى المجالس الرومانسية-31 إلى أرجينين يوفر بقايا الحفازة اللازمة في
رابطة الدول المستقلة، وبالتالي يزيد من نشاط GTPase من ARL6. بدلا من ذلك، تحور منتدى المجالس الرومانسية-31 إلى الميثيونين قد تعيق ملزم من المغنيسيوم 2+ وγ-الفوسفات بسبب طبيعة مسعور من سلسلة الجانب الميثيونين. تم العثور على طفرة من بقايا الغليسين-169 إلى ألانين في بارديه-بيدل متلازمة 3 مرضى. بقايا الغليسين-169 تقع ضمن مربع G5 الحفظ (بقايا 162-165)، الذي يربط ليس فقط في حلقة جوانين GTP مباشرة ولكن أيضا تشكل شبكة الهيدروجين الترابط مع مخلفات مربع G4 والمخلفات التالية مربع G5. وتساهم هذه السندات الهيدروجين لتحقيق الاستقرار في النوكليوتيدات المربوطة
(الشكل 2 B). ومن المثير للاهتمام، وبقايا الغليسين-169 يساهم في هذه الشبكة الرابطة الهيدروجينية، وتشكيل روابط هيدروجينية مع حمض الغلوتاميك-172 والغليسين-173
(الشكل 2 B). وأخيرا، تم العثور على بقايا لوي-170 لتكون جزءا من نواة مسعور مع سلاسل جانب إيل-33 ولوي-37
(الشكل 2 C). سوف تحور في بقايا لوي-170 إلى التربتوفان المرجح يعطل التعبئة والتغليف لهذه النواة مسعور وزعزعة استقرار البروتين.
ولذلك، التحليلات البنيوية لدينا ويتوقع أن الثريونين-31 الطفرات من المحتمل أن تؤثر على وجه التحديد النوكليوتيدات ملزمة، في حين توقع الغليسين-169 ولوي-170 الطفرات تؤثر في المقام الأول طي البروتين، مما أدى إلى عدم القدرة على ربط النوكليوتيدات والتحلل بواسطة الخلية الآلات. علاوة على ذلك، توقع كل الطفرات أن يؤدي إلى خسارة من وظيفة.
للتحقق من صحة تجريبيا البيانات البلورات، أجرينا فحوصات ملزمة النوكليوتيدات على البروتينات المؤتلف ARL6. تم اختبار من النوع البري ومتحولة البروتينات ARL6 الموسومة GST في المقايسات نقطة وصمة عار للربط إلى [α- 32 P] GTP، وذلك باستخدام GST وحدها كوسيلة لمراقبة سلبية. كما هو الحال مع GST وحده، وARL6 الموسومة GST (T31M) وARL6 (G169A) المسوخ عرض ضئيلة أو معدومة ملزمة بالمقارنة مع من النوع البري ARL6، في حين أن ARL6 (T31R) متحولة ملزمة مستويات أعلى بكثير من النوكليوتيدات
(التكميلي الشكل 1 ، A-C). على الرغم من أن [α- 32 P] تم استخدام GTP، وكمية صغيرة من [α- 32 P] الناتج المحلي الإجمالي موجود في قارورة المصدر. لذلك، لتحديد طبيعة النوكليوتيدات المربوطة، قدمنا أيضا المتغيرات الموسومة ب [γ- 32 P] GTP، وفي هذه الحالة فقط من النوع البري ARL6 عرض النشاط ملزم النوكليوتيدات فوق ضبط GST حدها
(التكميلي الشكل 1 D ). على الرغم من أن هذه النتائج قابلة للمقارنة مع تلك التي نشرت مؤخرا من قبل كوباياشي
وآخرون (57)، الذي كان FLAG- أو البروتينات الموسومة GST، فمن المعروف أن GTPases الصغيرة غالبا ما تكون حساسة لإضافة الترميز المنطقة في اجتماعهم N أو C محطة و أن هذا يمكن أن تتداخل مع الاستقرار و / أو وظائفهم. لهذا السبب، نحن كما نفذت الدراسات التي تستخدم إصدارات لازال من النوع البري والبروتينات ARL6 متحولة
(الشكل 3، A
و B). وعلى غرار النتائج المقدمة أعلاه، ARL6 (T31M)، ARL6 (G169A)، وARL6 (L170W) المسوخ أظهرت ضئيلة أو معدومة ملزمة من النوكليوتيدات الموجودة في [α- 32 P] خليط GTP، في مقابل ARL6 (T31R) ، الذي عرض تقارب أعلى بكثير لالنوكليوتيدات
(الشكل 3، C
وF). وبعد ذلك تستخدم TLC لتحديد طبيعة النوكليوتيدات المسمى بد أن ARL6. أظهرت النتائج أنه على الرغم من النوع البري ARL6 لا بد في المقام الأول إلى GTP، ARL6 (T31R) يبدو المنضم بدقة إلى الناتج المحلي الإجمالي
(الشكل 3 D). وبالتالي تظهر لدينا البلورات والبيوكيميائية النتائج مجتمعة أن كل من الخيارين ARL6 المسببة للأمراض يفترض، في الشكل الذي سيكون موجودا في المرضى
(أي غير محدد)، عرض ألغى النوكليوتيدات ملزم، مع ضئيلة أو معدومة ملزمة في حالة ARL6 (T31M)، ARL6 (G169A)، وARL6 (L170W) وعلى سبيل الحصر في الناتج المحلي الإجمالي في حالة ARL6 (T31R)
(الشكل 3 F).

الشكل 3.
BBS3 المتغيرات المسببة للأمراض تمتلك تلغى الأنشطة ملزمة النوكليوتيدات. A، Coomassie الملطخة جل SDS يظهر تحريض GST وحدها، غير محدد النوع البري ARL6، وT31M، T31R، وG169A المتغيرات ARL6. B، Coomassie الملون جل SDS-بولكرلميد يظهر على وقد رصدت نسبة من إجمالي المحللة خلية (T) التي لا تزال في جزء قابل للذوبان (S) لعينات من A بعد 3-ح الاستقراء. C، قسامة من الكسور القابلة للذوبان من B على نتروسليلوز وبحث مع [α- 32 P] GTP. GST، سيطرة سلبية، لا يلزم النوكليوتيدات، في حين ARL6 يربط. ARL6 (T31R) يربط أكثر بكثير من النوع البري ARL6، وT31M وG169A المتغيرات ربط ضعيف جدا. D، TLC تبين طبيعة النوكليوتيدات رديولبلد بد أن للذوبان ARL6 أو البديل أشكال ازال من ARL6. من النوع البري (غير محدد) ARL6 يربط كل من الناتج المحلي الإجمالي وGTP، ولكن المسوخ لازال (T31M وT31R) ربط أساسا الناتج المحلي الإجمالي. E، Coomassie الملطخة SDS هلام تظهر الكسور بيليه من النوع البري والبديل أشكال (T31M، T31R، G169A، L170W ) من البروتين ARL6، بفعل مماثل إلى A في تجربة منفصلة. F، كما هو الحال مع E إلا أنه بعد SDS-PAGE تم تحويل البروتين إلى غشاء النيتروسليلوز، سمح لإعادة أضعاف، وبحث مع [α- 32 P] GTP .
حتى الآن، تم العثور على جميع البروتينات BBS الثدييات درس في توطين إما في الجسيم المركزي / الجسم القاعدية أو في الأقمار الصناعية مريكزي المجاورة، التي تمثل مواقع رئيسية للبروتينات ذات دور الهدبية. على وجه التحديد، BBS2، BBS5، وBBS13 / MKS1 شارك في توطين مع الجسيم المركزي / القاعدية علامة الجسم γ تويولين
(38، 58، -، 60)؛ BBS4، BBS8، وBBS14 / CEP290 شارك في توطين مع PCM1 في الأقمار الصناعية مريكزي
(61، -، 63)؛ ويوجد BBS6 داخل أنبوب المحيطة centrioles المادية محيط بالمريكز
(64). لدهشتنا، واثنين من الدراسات السابقة المتمركزة الأخضر الفلورسنت ARL6 البروتين الموسومة وهيماغلوتينين الموسومة في جولجي
(65) أو في العصارة الخلوية
(66) من خلايا nonciliated، على التوالي. لذا سعينا لتحديد توطين التحت خلوية من ARL6 الذاتية في خط خلايا مهدبة.
لإجراء تجارب مناعية لدينا، نحن العاملين الأجسام المضادة الموجهة ضد N- والحواتم C-الطرفية من ARL6 الإنسان (المسمى ARL6N وARL6C، على التوالي). من خلال تحليل لطخة غربية، سواء الاعتراف ARL6 لكن ليس البروتين ARF6 ترتبط ارتباطا وثيقا، وتستخدم كعنصر تحكم للخصوصية
(الشكل التكميلي. 2).في الماوس النخاع جمع الخلايا الظهارية اصق (IMCD3)، وهما أضداد ARL6 وصمة عار على وجه التحديد نقاط وهما شبه النووية التي شاركت في توطين مع علامة centrosomal γ تويولين
(الشكل 4،
A وB). ويمكن ملاحظة إشارة ARL6 في centrosomes طوال دورة الخلية، مما يشير إلى وجود ارتباط محددة مع ديناميكية مركز أنيبيب المنظمة. في الطور البيني الخلايا، ارتبط ARL6 مع الجسم القاعدية، الذي الرياضية هدب أن يتم الكشف عن استخدام أجسام مضادة ضد تويولين الأسيتيل
(الشكل 4 B). أكدنا على centrosomal / القاعدية توطين جثة ARL6 في خط خلايا مهدبة الثاني، الخلايا الظهارية الصباغية وهي الشبكية (hTERT-RPE) (الشكل 5). الأهم من ذلك، نحن أيضا لاحظ شارك في توطين ARL6 مع الهيئات القاعدية في عدة أقسام أنسجة الثدييات مهدبة، بما في ذلك الخصية والمخ والكلى
(التكميلي الشكل 3، A-C). تظهر هذه البيانات التي الزميلة ARL6 / BBS3 مع الهياكل مريكزي / الهيئات القاعدية في مجموعة متنوعة من الخلايا المهدبة
في الجسم الحي.

الشكل (4).
ARL6 هو حسن النية / القاعدية البروتين المرتبط الجسم centrosomal أن يموضع بشكل مستقل عن الشبكة أنيبيب أو المحرك الجزيئي داينين. A، ARL6 الذاتية، لاحظ باستخدام الأجسام المضادة ARL6C (الحمراء) في الخلايا IMCD3 مهدبة، لوحظ في نقاط وشبه النووية أن شارك في توطين مع γ تويولين (الخضراء)، وcentrosomal / القاعدية علامة الجسم (الصفراء في الصورة المدمجة). أقحم، 2 × التكبير من المنطقة الموضحة. اتسخت DNA الأزرق مع 4، 6-diamidino-2-phenylindole. النصال أشر إلى ARL6 توطين لcentrosomes. B، ARL6 الذاتية (تصور باستخدام الأجسام المضادة ARL6N في الخلايا IMCD3؛ أحمر) لا يزال يرتبط مع الجسيم المركزي (ملحوظ باستخدام الأجسام المضادة ضد γ تويولين؛ الأخضر) في جميع مراحل دورة الخلية (الصفراء إشارة في الصورة المدمجة). لاحظ أن الخلية البيني ومهدبة (ملطخة الأجسام المضادة ضد الأسيتيل (ACET) تويولين؛ الأخضر). C والعلاج نوكودازول، ولكن ليس العلاج التحكم (DMSO)، إما 1 ساعة أو بين عشية وضحاها (O / N)، غير كافية ل تدمير شبكة أنيبيب في الخلايا IMCD3 يعاير كل α تويولين تلطيخ (الأخضر)، لكنه لا يملك أي تأثير على ARL6 (الأحمر) الترجمة. النصال أشر إلى ARL6 المترجمة إلى centrosomes في الخلايا تعامل مع بين عشية وضحاها نوكودازول. D، وتثبيط المحرك الجزيئي ويتحقق داينين من خلال overexpression من الأخضر الفلورسنت الموسومة البروتين P50-dynamitin (الأخضر) الذي أبداه القدرة على mislocalize PCM1 (اللوحة اليمنى)، وهو بروتين المعروف أن تعتمد على داينين للتوطين في centrosomal (لاحظ تلطيخ المشتتين في الخلايا معربا P50-dynamitin مقارنة مع الخلايا لا تعبر عن المانع داينين). ARL6 (اللوحة اليسرى لم يتم mislocalized) عندما هى overexpressed P50 dynamitin. النصال تشير إلى توطين السليم للARL6 في الخلايا معربا عن P50-dynamitin.

الشكل 5.
ARL6 يموضع في النهاية البعيدة من الجسم القاعدية في نمط على شكل حلقة. A، مناعية باستخدام الأجسام المضادة ARL6C يظهر ARL6 توطين إلى النهاية البعيدة من centrioles الأم والهيئات القاعدية في الخلايا hTERT-RPE. أقحم، صورة مكبرة للاختيار المنطقة. B والصور المجهري متحد البؤر باستخدام الأجسام المضادة ARL6N في الخلايا hTERT-RPE يدل على أن ARL6 يموضع في نمط يشبه عصابة، تماما البعيدة إلى centrioles الأم، في الخلايا nonciliated والمهدبة.
على غرار BBS6
(64)، ولكن على عكس BBS4 ومن البروتين التفاعل PCM1
(63)، وتوطين ARL6 إلى الجسيم المركزي / الجسم القاعدية هو-أنيبيب مستقل، وعلاج الخلايا IMCD3 مع أنيبيب depolymerizing نوكودازول المخدرات لا يؤثر هذا التعريب
(الشكل 4 C). وعلاوة على ذلك، فإن توطين ARL6 هو أيضا يتأثر في الخلايا حيث تعطلت داينين السيارات إلى الوراء من قبل overexpression من P50-dynamitin
(الشكل 4 D). معا، وهذه النتائج تدعم بقوة فكرة أن ARL6 الذاتية / البروتين BBS3 هو
حسن النية البروتين المرتبط مريكز التي هي موجودة في أنيبيب تنظيم مركز في تقسيم الخلايا وفي الجسم القاعدية في الخلايا المهدبة، في microtubule- وdynein- بطريقة مستقلة.
بهدف تحديد بدقة موقف ARL6 داخل الجسم القاعدية، أجرينا التصوير متحد البؤر على الخلايا hTERT-RPE ملطخة الأجسام المضادة على حد سواء المضادة للARL6. ومن المثير للاهتمام، تم العثور على بروتين ARL6 إلى التركز في نهاية واحدة من الهيكل القائم على أنيبيب. على وجه التحديد، والخلايا البيني تفتقر إلى هدب في وقت تظهر تثبيت ARL6 توطين في نمط تشبه الحلبة في نهاية البعيدة للمريكز واحد فقط، من المفترض ان مريكز الأم. للخلايا التي لا تمتلك هدب، يتم وضع حلقة من ARL6 مماثل، وتحديدا عند تقاطع بين الجسم القاعدية والخيط المحوري الهدبية
(الشكل 5،
A وB). هذه النتائج تشير إلى أن ARL6، وهي ليست موجودة في مجمع BBSome
(37)، وتعمل ضمن منطقة متميزة وفريدة من نوعها من الجسم القاعدية مقارنة مع جميع البروتينات BBS مدروسة الأخرى. توطين موضوعه متفقا مع كونها قريبة أو في الألياف الانتقالية، التي تشكل جزءا من ما يسمى بوابة الهدبية.
اقترح توطين فريد من ARL6 الثدييات / BBS3 في قاعدة أهداب أن هذا GTPase صغيرة تمتلك دورا في تشكيل هدب في خلايا الثدييات، وربما عن طريق الاتجار الغشاء الهدبي. لتحقيق هذا الاحتمال، اتخذنا الاستفادة من حقيقة أن الإفراط في GTPase صغير أشكال متحولة غالبا ما تستخدم لتحقيقاتها
في الجسم الحي وظائف (67). مصادفة، والطفرات المقابلة لبقايا الثريونين-31، التي وجدناها سابقا في ولاية متماثل في المرضى الذين يعانون BBS
(28) ويؤدي إلى بروتين غير الساحلية الناتج المحلي الإجمالي
(الشكل 3 D)، ويشيع استخدام كأشكال السلبية المهيمنة. ولذلك نحن على استعداد لبناء التعبير الثدييات لARL6 (T31R)، وكذلك تحمل طفرة آخر واحد من الأحماض الأمينية في GLN-73، وتوقع أن تسفر عن والشكل غير الساحلية GTP نشط بشكل جوهري من البروتين (68). وإن كان في معظم الحالات أشكال غير الساحلية GTP خلق GTPases صغيرة نشطة بشكل جوهري، في بعض الحالات يبدو أن هذا هو ركوب الدراجات من النوكليوتيدات ما هو ضروري من أجل وظيفة البروتين. في تلك الحالات، وشكل غير الساحلية GTP يتصرف أشبه سلبية المهيمنة، يشبه إلى حد كبير شكل غير الساحلية الناتج المحلي الإجمالي، كما هو الحال بالنسبة لشكل غير الساحلية GTP من ARL6 (انظر أدناه). ومن المثير للاهتمام، وشكل غير الساحلية GTP من ARF6 (Q67L)، والذي يحدث أيضا أن تكون GTPase صغيرة المنتدى الاقليمى للاسيان وفقا لأعلى درجة من الهوية إلى ARL6، يعرض أيضا هذه الخاصية (69، -، 71).
الإفراط في البرية من نوع ARL6، أو أي من المتغيرات، لم ARL6 (T31R) أو ARL6 (Q73L)، في الخلايا hTERT-RPE لا يؤدي إلى تغييرات ملموسة على الانقسام، أنيبيب الهيكل الخلوي، أو مورفولوجيا الخلايا مقارنة بالمجموعة الضابطة ( لا تظهر البيانات). ومع ذلك، باستخدام تويولين الأسيتيل كعلامة للأهداب، لاحظنا نسبة أعلى بكثير من الخلايا التي تملك أهداب في تلك مع transfected إما ARL6 (T31R) أو ARL6 (Q73L) بنيات overexpression بالمقارنة مع الخلايا السيطرة (41.5 ± 3.5 و 54.5 ± 6.5٪
مقابل 25 ± 4٪،
ع <0.001 مقارنة مع الشاهد)
(الشكل 6. A). ومن اللافت للنظر، تم الكشف عن النمط الظاهري المعاكس، وهي نسبة أقل دلالة إحصائية الخلايا مع أهداب، في تلك مع transfected من النوع البري ARL6 بناء (9 ± 2
مقابل 25 ± 4٪،
ع <0.001 مقارنة مع الشاهد)
(الشكل 6. A). كان النمط الظاهري آخر لاحظت فرقا كبيرا في طول الأهداب في الخلايا معربا عن أشكال متحولة من ARL6. في حين كان متوسط طول أهداب من الخلايا المصابة بالعدوى بالنقل مع من النوع البري ARL6 لا يختلف كثيرا عن السيطرة، وأهداب من ARL6 (T31R) - كانت وARL6 (Q73L) خلايا -transfected لفترة أطول كثيرا (4.79 ± 0.19 و 5.21 ± 0.36 ميكرون
مقابل 2.4 ± 0.09 ميكرومتر،
ص <0.001)
(الشكل 6. B). معا، وتشير هذه النتائج إلى أن BBS3 يعمل على تعديل وجود وبنية أهداب الابتدائي؛ من النوع البري النتائج BBS3 الزائدة في الخلايا المهدبة أقل، في حين أن إلغاء وظيفتها يؤدي إلى نسبة أكبر من الخلايا المهدبة مع أهداب طويلة.

الشكل 6.
overexpression من النوع البري ARL6 / BBS3 والمتغيرات النتائج في الشذوذ الهدبية. و transfected الخلايا hTERT-RPE مع واحدة من ثلاث بنيات، pCMV-BBS3 (overexpressing من النوع البري غير محدد وBBS3)، pCMV-T31R، أو pCMV-Q73L (overexpressing المتغيرات ازال). وتم قياس كمية الظواهر الناتجة أيام 2 بعد ترنسفكأيشن. A، تم تحديد نسبة الخلايا التي تمتلك هدب عن طريق عد عشوائيا> 100 خلية من كل السكان. وأشرطة الخطأ تمثل الخطأ المعياري بين تجارب مكررة. B، تم تحديد طول هدب عن طريق قياس> 30 أهداب عشوائي من السكان. وأشرطة الخطأ تمثل الخطأ المعياري بين تجارب مكررة. النجمة تمثل فروق ذات دلالة إحصائية مقارنة مع الشاهد. C، صور تمثيلية تظهر الاختلافات في طول هدب بين الخلايا hTERT-RPE التي كانت وهمية transfected (السيطرة) أو transfected للتعبير عن إصدارات لازال من النوع البري ARL6، ARL6 (T31R)، أو ARL6 (Q73L) المتغيرات. دابي، 4، 6-diamidino-2-phenylindole.
اكتشاف أن أهداب هي المركزية لمختلف مسارات الإشارات قد عززت بشكل كبير من فهمنا لدور هذه العضيات في التنمية والمرض. كجزء من وظيفتها في الإشارات بين الخلايا، وهدب يقمع بشكل طبيعي، β-تعتمد على catenin إشارات WNT الكنسي (72،
73) ومطلوب لnoncanonical، β-catenin مستقل يشير WNT. فقدان العديد من البروتينات BBS، بما في ذلك BBS2، BBS4، وBBS6، يؤدي إلى زيادة بيتا catenin النشاط النسخي عندما تحفزها إشارة Wnt3a الكنسي
(73)، في حين لم يقم overexpression من BBS4 تأثير العلني على النشاط β-catenin.
5
لاستكشاف الدور المحتمل للARL6 / BBS3 في إشارات Wnt، عبرنا عن ARL6 / BBS3 في خط الخلية hTERT-RPE1 مع transfected مراسل البلازميد pTOPflash
(الشكل 7) وخط HEK293T مستقر خلية مراسل WNT (التكميلي الشكل 4 A ). ومن المثير للاهتمام، وتأثير ARL6 / BBS3 على النشاط β-catenin هو تعتمد على الجرعة، لأن overexpression يؤدي إلى التقريبية 2- أو 6 أضعاف زيادة استجابة Wnt3a في hTERT-RPE1
(الشكل 7) وHEK293T (التكميلي الشكل 4 A) خط الخلية مقارنة مع خلايا من النوع البري، على التوالي. في المقابل، overexpression من ARL6 (T31R) وARL6 (Q73L)، التي تعرف الآن أن تسبب التشوهات الهدبية
(الشكل 6)، لم يكن لديها تأثير على β-catenin النشاط النسخي (الشكل 7). على الرغم من ARL6 (T31R) يمثل طفرة المسببة للأمراض، ARL6 (Q73L) ليست طفرة المحددة في المرضى BBS3. لذلك، لمواصلة التحقيق أم لا يشير WNT قد تلغى في المرضى الذين يعانون BBS، اختبرنا ردا على Wnt3a يجند في متغيرات إضافية ARL6 overexpressed المسببة للأمراض، وهي ARL6 (G169A) وARL6 (L170W). وعلى غرار ARL6 (T31R) وARL6 (Q73L)، لاحظنا فقدان responsivity للإشارة Wnt3a وβ-catenin النشاط النسخي مقارنة عندما هى overexpressed من النوع البري ARL6
(التكميلي الشكل 4 B). معا، تظهر هذه النتائج أن ARL6 / BBS3 يمكن أن تعدل إشارات Wnt في خلايا الثدييات والتي قد يكون لها تأثير مختلف عن البروتينات BBS الأخرى (انظر أدناه).

الشكل 7.
overexpression من النوع البري ولكن ليس متحولة أشكال ARL6 / BBS3 يغير الاستجابة لWnt3a يجند. في الخلايا مراسل hTERT-RPE TOPflash، overexpression من ARL6 / BBS3 يؤدي إلى زيادة النشاط β-catenin ردا على Wnt3a يجند ولكن لا ديسريغولاتيون العالمي للβ يشير -catenin. في المقابل، overexpression من ARL6 (T31R) وARL6 (Q73L) ليس له تأثير ملحوظ على β-catenin النشاط النسخي. الخلايا باستمرار 70-80٪ متكدسة ومهدب كما لوحظ من قبل تلطيخ مع مضاد للالأسيتيل تويولين الأضداد (لا تظهر البيانات). أشرطة الخطأ تمثل الانحراف المعياري. فقط النوع البري (BBS3 WT عينات) تختلف إحصائيا من مكافحة ناقلات الفارغة (ع <0.05).
هنا، نقدم التركيب البلوري للARL6 / BBS3، أول المعلومات الهيكلية عالية الدقة لبروتين BBS أو IFT. لأن GTPases المنتدى الاقليمى للاسيان وعادة ما يكون تقارب أعلى للناتج المحلي الإجمالي من GTP (على سبيل المثال ARL5A، رموز PDB
2H16 /
2H17؛ ARL8A، رموز PDB 2H18 /
1ZD9؛ ARL8B، رمز PDB
2AL7؛ وSAR1A، PDB كود
2GAO)، البيانات الهيكلية لدينا على ARL6 كانت غير متوقعة، خاصة وأننا كنا قادرين على حل هيكلها في ولاية محددة GTP (74). أحد الأسباب المحتملة لهذا العقار تبدو فريدة من ARL6 هو أن منطقتها التبديل II تفضل التشكل محددة GTP بسبب الرابطة الهيدروجينية بين سلسلة الجانب سر-71، والتي هي فريدة من نوعها بين GTPases صغيرة / ARF مثل المنتدى الاقليمى للاسيان، وأميد مجموعة من GLN-73
(الشكل 2 A). وبالتالي، قد يكون ARL6 أعلى النسب للGTP من الناتج المحلي الإجمالي، وهو الاكتشاف الذي يتم اعتماد كيميائيا
(الشكل 3 والشكل التكميلي 1). بالإضافة إلى ذلك، فمن المتوقع لليسين الغنية N-14 محطة بقايا ARL6، لم يكن موجودا في كياننا لتشكيل مسعور α-الحلزون بدلا من الحلزون متقابلة الزمر، كما هو الحال مع غيرها من GTPases الصغيرة الأسرة المنتدى الاقليمى للاسيان. ومن المتوقع myristoylation من بقايا الغليسين-2 في ARL6 أيضا أن يكون من غير المحتمل (32). وبالتالي، فمن الممكن أن كلا من النوكليوتيدات خصائص الملزمة ARL6، وكذلك الطريقة التي الغشاء ينظم ملزمة، قد تكون مختلفة عن تلك التي أفراد الأسرة الآخرين.
الأهم من ذلك أن بيانات التركيب البلوري شرح تأثير الطفرات المسببة للأمراض قد أن على البروتين ARL6 / BBS3. هذه الطفرات إما تؤثر بقايا الحرجة اللازمة لGTP ملزم،
أي الثريونين-31، أو أنها تؤثر على روابط هيدروجينية ضمجزيئي في مثل هذه الطريقة ربما لسبب misfolding المحلية / العالمية كذلك،
أي الغليسين-169 ولوي-170. لدينا دراسات وظيفية تتفق مع هذه الفكرة. وARL6 (T31M)، ARL6 (G169A)، وARL6 (L170W) المتغيرات عرض ضئيلة أو معدومة النوكليوتيدات ملزمة، في حين لوحظ ARL6 (T31R) البديل في حالة غير الساحلية الناتج المحلي الإجمالي تختلف عن نظيرتها من النوع البري، الذي كان أساسا GTP محددة
(الشكل 3). في ظل هذه الظروف، ومن المتوقع أنشطة GTPase صغير قد انخفضت، وهو احتمال، كما هو مذكور أدناه، أكدنا من خلال مراقبة الهدبية ويشير الظواهر.
وتوضح النتائج التي توصلنا إليها أن ARL6 / BBS3 وجدت في نمط تشبه الحلبة في النهاية البعيدة من الجسم القاعدية ومعظم المنطقة القريبة من الخيط المحوري الهدبية
(الشكل 5). هذا التعريب الجسم القاعدية للARL6 / BBS3 يتسق مع النتائج السابقة للبروتينات BBS الأخرى (38، 58، 60، -، 63). ومع ذلك، فإن الموقف الدقيق للARL6 / BBS3 في النهاية البعيدة من الهيئات القاعدية هي فريدة من نوعها بين البروتينات BBS لكن مماثلة لتلك التي ذكرت لالتهاب الشبكية الصباغي 2 (RP2) نديد في
C. ايليجانس (34) والمثقبيات البروسية (75)، والتي تشكل أيضا هياكل على شكل حلقة في قاعدة الأهداب. ومن المثير للاهتمام، ومن المعروف RP2 للتفاعل معها، ويكون بمثابة-المنشط GTPase ل، وGTPase ARL3 صغيرة
(76)، وظيفتها في تشكيل هدب تم توثيقه في الكائنات الحية المختلفة (33، -، 35).
الملاحظ أيضا هو حقيقة أن الموقع التحت خلوية عمل ARL6 إما في، أو على مقربة، والألياف الانتقالية في النهاية البعيدة من جميع الهيئات القاعدية. نصائح من هذه الألياف، التي تنضم إلى الغشاء الهدبي، يعتقد أنها مواقع لرسو السفن لجزيئات IFT (19، 22). وبالنظر إلى أن
C. ايليجانس ARL-6 / BBS-3، مثل كل اختبار آخر
C. ايليجانس البروتينات BBS (BBS-1، -2، -5، -7، و-8) (77،
78)، وشركاء مع الآلات IFT (أي تحركات بطريقة تشبه IFT في أهداب الحسية)، ونحن التكهن بأن ARL6 / BBS3 قد تتفاعل مع البروتينات BBS الأخرى والآلات IFT قرب أو في هذه المنطقة الألياف الانتقالية. تساعد الألياف أيضا تشكيل بوابة الهدبية، مما يحد من دخول الحويصلات في الفضاء الهدبي، مثل أن حويصلات جولجي المستمدة من تحمل قفص الاتهام البضائع الهدبي والصمامات على مقربة من الألياف (21، 22). لذلك، نظرا للدور أنشئت للبروتينات BBS في الاتجار داخل خلايا الجسم، وIFT في أهداب، يمكن ARL6 / BBS3 تسهيل تصور / تعدل الانتقال بين حويصلي والاتجار intraciliary. سيكون تحديا من نوع خاص لتشريح وظائف محددة من ARL6 / BBS3، باعتبارها واحدة سوف تحتاج إلى أن تأخذ في الاعتبار قبل القاعدية الجسم / الهدبية الاتجار غشاء فضلا عن الاتجار intraciliary (IFT) عمليات -associated. على الأرجح السابق يتضمن العديد من GTPases الصغيرة، بما في ذلك RAB8، RAB17، وRAB23، وربما لمجموعة فرعية من البروتينات (التي تحتوي على V
X PX الزخارف)، ARF4 وRAB11 (37، 79، 80)؛ يعتمد هذا الأخير على مجموعة من اللاعبين الناشئة، بما في ذلك البروتينات RAB IFT25، RAB8، والإفتاء-2، فضلا عن ARL13B (34، 36، 37، 39).
من خلال فائض الانتاج في الخلايا المهدبة كلا من النوع البري ARL6 وشكلين متحولة من GTPase صغير ARL6، وكشف لنا دور محتمل لهذا GTPase صغيرة في هدب التفكيك. على وجه التحديد، overexpression من النوع البري النتائج ARL6 إلى انخفاض في تشكيل هدب، لاحظ النمط الظاهري أيضا على الصغيرة بالتدخل RNA ضربة قاضية من BBS3 في مسح لجميع
BBS الجينات (80)؛ من ناحية أخرى، overexpression من ARL6 (T31R) وARL6 (Q73L)، التي تمنع نشاط ARL6 الذاتية، يسبب زيادة في تشكيل هدب وطول
(الشكل 6).
من هذه النتائج، فإننا نقترح أن ARL6 قد تسهل، يحتمل أن تكون من خلال تهريب بعض البضائع الهدبية، وارتشاف من الأهداب. مطلوب هذه العملية لتحرير centrioles (الجسم القاعدية)، لاستخدامها كمراكز أنيبيب المنظمة، قبل دورة الخلية إعادة دخول (81). على الرغم من أن هذه وظيفة ستكون الأولى من نوعها البروتينات BBS، البروتينات IFT هي ثيقا المعنية في ارتشاف الهدبية (17).
ومن المثير للاهتمام، الإفراط في ما يعادل (يفترض أن الناتج المحلي الإجمالي مقفلة) أشكال متحولة من ARL6 (T31R)
(الشكل 6 B) وRAB8 (T22N)
(37) وتنتج عيوب الهدبية المعاكس أساسا، مشيرا إلى وظائف معارضة لاثنين من GTPases الصغيرة. ومن المحتمل ذات الصلة التي في الشاشة الزرد عن التفاعلات الوراثية بين
BBS الجينات،
ARL6 تم العثور على التفاعل مع
BBS1 (82)، الذين المنتج الزميلة جسديا مع RAB8 (الجين 37). على العموم، تشير هذه البيانات إلى جمعيات الوظيفية المحتملة بين BBSome، ARL6 / BBS3، وRAB8 في التوسط تكون الأهداب وهدب ارتشاف. الآن هناك علاقة بين BBSome والهدبية ارتشاف. وقد تبين أن البروتين BBIP10 BBSome المصاحب للتفاعل مع HDAC6، الذي أستلة أنيبيب النشاط يؤثر الهدبية التفكيك (83، 84). بالإضافة إلى ذلك، وتورط البروتينات BBS أيضا في إزالة البروتينات من أهداب في
كلاميدوموناس (100)، بما يتفق مع دور محتمل في ارتشاف.
بدلا من ذلك، يمكن أن BBS3 المشاركة في هدب السيطرة طول، وهي عملية نعرف أن ينظم بإحكام في
كلاميدوموناس reinhardtii. عندما
كلاميدوموناس يتم تحفيز الخلايا لاستئصال الآفة، ويتم إجراء بدلاء لهم إلى ما قبل deflagellation أطوال في أقل من 2 ساعة (85). وعلاوة على ذلك، عندما يتم بتر السوط واحد فقط، فإن السوط المتبقية تقصير حتى يصل نظيرتها بنفس الطول، في الوقت الذي كل من سياط تنمو إلى طول محدد مسبقا من (85). لا يقل عن سبعة البروتينات، المشفرة بواسطة أربعة سياط طويلة
(LF) وثلاثة أسواط (قصيرة الموجات الدقيقة) الجينات، وتنظيم هذه العملية في
C. reinhardtii (86، -، 89). يتم وضع BBS3 من الناحية المثالية، عند أو بالقرب من بوابة الهدبية، لتعمل في مراقبة طول الهدبية. على وجه التحديد، فقد تم الافتراض أن الألياف الانتقالية والآلات الموجودة في هذه المنطقة قد فرض رقابة صارمة على حركة المكونات داخل وخارج المقصورة الهدبية، على سبيل المثال البروتينات IFT، بطريقة مماثلة لتلك التي من مجمع المسام النووي (19) .
فقدان البروتينات BBS، بما في ذلك BBS1، BBS4، وBBS6، يؤدي إلى استجابة تضاف إلى إشارات الكنسي WNT مثل Wnt3a مقاسا β-catenin النشاط النسخي (73). وبالنظر إلى هذه النتائج، فإنه لم يكن متوقعا أن overexpression، وليس ضربة قاضية، من بروتين آخر BBS، وهي BBS3، أدى إلى رد فعل المعزز مماثلة لWnt3a يجند الكنسي
(الشكل 7). نقترح أن زيادة النشاط β-catenin ينظر في البرية من نوع الخلايا overexpressing BBS3 مرتبط نسبتهم أقل من أهداب
(الشكل 6 A). وقد اقترح أن أهداب تؤثر على التوازن بين الإشارات WNT الكنسي وnoncanonical داخل الخلية من خلال تفضيل المسار noncanonical (73)؛ وبالتالي، زيادة التفكيك الأهداب في الخلايا overexpressing من النوع البري BBS3 يفضلون يشير WNT الكنسي، كما لاحظنا.
على الرغم من أن قد يتوقع المرء أن يرى أثر عكسي، انخفضت هما النشاط بيتا catenin في الخلايا overexpressing وARL6 (T31R) وARL6 (Q73L) المتغيرات، التي تبين أيضا هنا أن تسفر عن الظواهر الهدبية، لاحظنا أي تأثير كبير على β -catenin النشاط النسخي من overexpression من هذه الطفرات السائدة
(الشكل 7). على الرغم من أن هذه النتيجة ليست بديهية على الفور، فإنه لا تتناسب مع دورنا المفترضة لBBS3، وهو ذلك الجزء من وظيفتها قد يكون للمساعدة في تهريب مكونات الغشاء الهدبي للخروج من مقصورة الهدبية. لتحقيق التوازن الديناميكي بين كل من WNT مسارات إشارات الكنسي وnoncanonical، والهدبية "التبديل" يجب أن تعمل بشكل صحيح، وأنه من المرجح أن يتطلب الاتجار الصحيح للمكونات الغشاء الهدبي ليس فقط في، ولكن أيضا من ومقصورة الهدبية. إذا مكونات الهدبية للمسار WNT
(على سبيل المثال inversin) (90، -،
92) و، لا يتم الاتجار بهم بشكل صحيح من هدب، ومن المرجح أن يكون ضعف التحول إلى إشارات noncanonical، والنوع البري مستويات النشاط β-catenin يكون لاحظت، كما هو الحال مع نتائجنا. قد يوحي هذا أن التحول الهدبية التي تفضل قد يكون noncanonical إشارات Wnt مختلة عند منسوخ BBS3، وهو احتمال يحتاج إلى مزيد من التحقيق.
على الرغم من أن نتائجنا تتفق مع النماذج الحالية تشير إلى دور للأهداب في إشارات Wnt، فإنه سيكون فقط المناسب أن نعترف أن شرط أهداب في كبح إشارات WNT الكنسي لا يزال مثيرا للجدل. ينبع الخلاف من النتائج التي تظهر نماذج الماوس IFT معيب ونماذج الزرد-الأهداب أقل تظهر من النوع البري من أنماط التعبير الكنسي WNT مكونات الإشارات وتستجيب عادة للالكنسي WNT يجند (93، 94). لكن، وكما هوانغ وشير
(94) الاعتراف، ضربة قاضية لهذه المكونات IFT النتائج حصرا في العيوب أهداب ويترك الجسم القاعدية أكثر أو أقل سليمة. لأن البروتينات BBS غالبا ما تكون القاعدية في المقام الأول، وفي بعض الحالات يشاركون في الاتجار العناصر الحاسمة centrosomal (المرتبط الجسم 61،
63)، فإنه يمكن أن تورط البروتينات BBS في إشارة WNT له علاقة مع دورها في الهيئات القاعدية، الأساس الذي بنيت أهداب. ولذلك، فإننا نؤكد أنه إذا لم أهداب ثم الهيئات القاعدية على أقل تقدير حاسمة لتقييد الكنسي إشارات Wnt من خلال التأثير على توازن ديناميكي بين الكنسي وnoncanonical إشارات Wnt.
ومن المثير للاهتمام، تعطلت التعبير عن Wnt3a في الخلايا العصبية في القلب قبل الهجرة قمة الزرد مع بوساطة morpholino ضربة قاضية من البروتين التفاعل ARL6 (ARL6IP) (95). وعلاوة على ذلك، فقد ARL6IP تحاكي العديد من الظواهر التي لوحظت في ضربة قاضية للبروتينات BBS في
دانيو rerio، وهي تشوهات القحفي، وعيوب في التنمية برعم الطرف، عيوب القلب، وتصبغ غير طبيعي، وتنكس الشبكية (82، 95، -، 97). وبالتالي يمكن أن طبيعة هذا التفاعل الوظيفي بين ARL6 وARL6IP يثبت أنه مفيد لفهمنا العمليات الخلوية والجزيئية التي تنظم وظيفة هدب، وارتباطه مع إشارات Wnt.
 « آخــر المشـاركــات »
« آخــر المشـاركــات »
 11-23-2015, 09:44 PM
11-23-2015, 09:44 PM












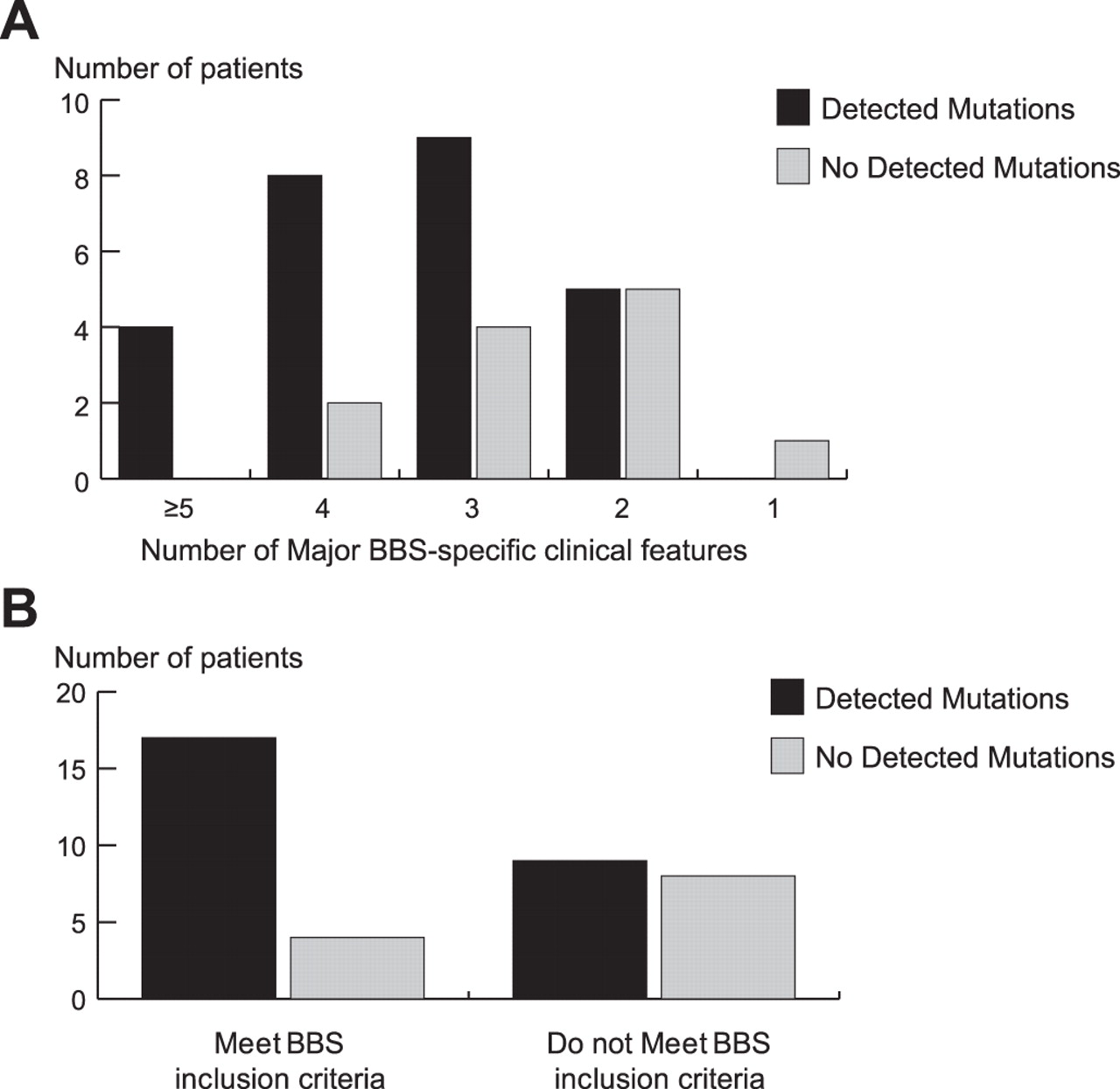
























 وترتبط 50 أعضاء معروفين من هذه العائلة إلى الأمراض التي تصيب البشر. باستخدام شاشة بيوينفورمتيك للجينات الهدبية في تركيبة مع التحليلات طفرية، حددنا ARL6 كما الجينات الكامنة بارديه-بيدل نوع متلازمة 3، وهو اضطراب multisystemic تتميز السمنة، والعمى، polydactyly، تشوهات الكلى وضعف الادراك
وترتبط 50 أعضاء معروفين من هذه العائلة إلى الأمراض التي تصيب البشر. باستخدام شاشة بيوينفورمتيك للجينات الهدبية في تركيبة مع التحليلات طفرية، حددنا ARL6 كما الجينات الكامنة بارديه-بيدل نوع متلازمة 3، وهو اضطراب multisystemic تتميز السمنة، والعمى، polydactyly، تشوهات الكلى وضعف الادراك  10 -6)؛ ثلاثة من هذه وقعت في الفترة الحرجة BBS3
10 -6)؛ ثلاثة من هذه وقعت في الفترة الحرجة BBS3 


 وتظهر S (برتقالي مظللة) باللون الأزرق. وتظهر التغييرات الأحماض الأمينية الناتجة عن الطفرات التي تم تحديدها في الأسر التي لديها BBS3 باللون الأحمر.
وتظهر S (برتقالي مظللة) باللون الأزرق. وتظهر التغييرات الأحماض الأمينية الناتجة عن الطفرات التي تم تحديدها في الأسر التي لديها BBS3 باللون الأحمر. 
 الوحيدات من heterotrimeric قناة البروتين استيراد SEC61 (المرجع
الوحيدات من heterotrimeric قناة البروتين استيراد SEC61 (المرجع  مللي ثانية -1 على التوالي. هذه المعدلات هي مماثلة لتلك التي من dynein- الآخر ويرتبط كينيسين البروتينات IFT
مللي ثانية -1 على التوالي. هذه المعدلات هي مماثلة لتلك التي من dynein- الآخر ويرتبط كينيسين البروتينات IFT 
 10 -6. لكل C. البروتين ايليجانس، جمعنا كبار H. ضرب العاقل من إخراج BLAST وفرز هذه وفقا لعدد الصبغيات والموقف. ثلاثة H. انخفضت مجموعات العاقل UniGene ضمن الفترة الحرجة BBS3
10 -6. لكل C. البروتين ايليجانس، جمعنا كبار H. ضرب العاقل من إخراج BLAST وفرز هذه وفقا لعدد الصبغيات والموقف. ثلاثة H. انخفضت مجموعات العاقل UniGene ضمن الفترة الحرجة BBS3  الشذوذ النظامية
الشذوذ النظامية  العرض العادي
العرض العادي

